Anelasmocephalus Simon, 1879
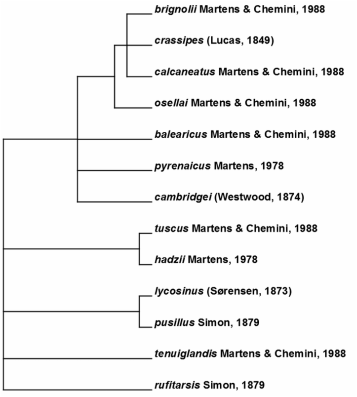
The tree is derived from the phylogenetic concepts of Martens and Chemini (1988) based on morphology.
Anelasmocephalus comprises small bodied (2.0-5.3 mm) soil- and litter-dwelling species, which are drop-shaped rather than dorso-ventrally flattened, in contrast to other Trogulidae. The head section is narrow, and the bifurcate head-cap is reduced to mere stubs covered with long papillae that replace the head above the chelicerae. Body and legs are more irregularly covered with papillae than in other Trogulidae and papillae are longer and set with long stout bristles. Species can be told apart upon tarsal formula, relation of length of tarsal segments, distribution and body size. The center of diversity is clearly Italy, where nine of the 13 described species occur.
Taxonomy and Discussion of Phylogenetic Relationships
The phylogenetic relationship of Anelasmocephalus within Trogulidae is not resolved but is probably closest to the morphologically similar Kofiniotis (Schönhofer and Martens 2010). Martens and Chemini (1988) organized the genus into species groups based on tarsal formula and genital morphology. It accords with a concept of closely related species being allopatric but members of different groups allowed sympatry. One exception is the species pair A. cambridgei and pyrenaicus, the latter having strongly derived male genitalia, probably to separate reproductively from the larger species A. cambridgei. Martens and Chemini (1988) pointed out differences between allopatric populations within several species (e.g. lycosinus, rufitarsis) that might indicate further cryptic differentiation.
Ecology and Life History
Most species of Anelasmocephalus inhabits well structured soil and litter layers in deciduous forest, often in combination with Fagus. Little is known about the biology of the single species. Pabst (1953) conducted detailed studies on A. cambridgei and showed them to prey on snails similar to Trogulus. Pabst found them to have a shorter generation time than Trogulus “nepaeformis”, which probably accounts for a faster mutation rate and longer branch lengths recorded in phylogenetic analyses of Trogulidae (Schönhofer and Martens 2010).
References
Martens, J. and C. Chemini. 1988. Die Gattung Anelasmocephalus Simon, 1879. Biogeographie, Artgrenzen und Biospezies-Konzept (Opiliones: Trogulidae). Zoologische Jahrbücher. Abteilung für Systematik, Geographie und Biologie der Tiere 115:1-48.
Pabst, W. 1953. Zur Biologie der mitteleuropäischen Troguliden. Zoologische Jahrbücher. Abteilung für Systematik, Geographie und Biologie der Tiere 82:1-46.
Schönhofer, A. L. and J. Martens. 2010. Hidden Mediterranean diversity: Assessing species taxa by molecular phylogeny within the opilionid family Trogulidae (Arachnida, Opiliones). Molecular Phylogenetics and Evolution 54 (1): 59-75.
Credits
Page created by Axel L. Schönhofer. Dave Carlson helped improve the English.
Anelasmocephalus comprises small bodied (2.0-5.3 mm) soil- and litter-dwelling species, which are drop-shaped rather than dorso-ventrally flattened, in contrast to other Trogulidae. The head section is narrow, and the bifurcate head-cap is reduced to mere stubs covered with long papillae that replace the head above the chelicerae. Body and legs are more irregularly covered with papillae than in other Trogulidae and papillae are longer and set with long stout bristles. Species can be told apart upon tarsal formula, relation of length of tarsal segments, distribution and body size. The center of diversity is clearly Italy, where nine of the 13 described species occur.
Taxonomy and Discussion of Phylogenetic Relationships
The phylogenetic relationship of Anelasmocephalus within Trogulidae is not resolved but is probably closest to the morphologically similar Kofiniotis (Schönhofer and Martens 2010). Martens and Chemini (1988) organized the genus into species groups based on tarsal formula and genital morphology. It accords with a concept of closely related species being allopatric but members of different groups allowed sympatry. One exception is the species pair A. cambridgei and pyrenaicus, the latter having strongly derived male genitalia, probably to separate reproductively from the larger species A. cambridgei. Martens and Chemini (1988) pointed out differences between allopatric populations within several species (e.g. lycosinus, rufitarsis) that might indicate further cryptic differentiation.
Ecology and Life History
Most species of Anelasmocephalus inhabits well structured soil and litter layers in deciduous forest, often in combination with Fagus. Little is known about the biology of the single species. Pabst (1953) conducted detailed studies on A. cambridgei and showed them to prey on snails similar to Trogulus. Pabst found them to have a shorter generation time than Trogulus “nepaeformis”, which probably accounts for a faster mutation rate and longer branch lengths recorded in phylogenetic analyses of Trogulidae (Schönhofer and Martens 2010).
References
Martens, J. and C. Chemini. 1988. Die Gattung Anelasmocephalus Simon, 1879. Biogeographie, Artgrenzen und Biospezies-Konzept (Opiliones: Trogulidae). Zoologische Jahrbücher. Abteilung für Systematik, Geographie und Biologie der Tiere 115:1-48.
Pabst, W. 1953. Zur Biologie der mitteleuropäischen Troguliden. Zoologische Jahrbücher. Abteilung für Systematik, Geographie und Biologie der Tiere 82:1-46.
Schönhofer, A. L. and J. Martens. 2010. Hidden Mediterranean diversity: Assessing species taxa by molecular phylogeny within the opilionid family Trogulidae (Arachnida, Opiliones). Molecular Phylogenetics and Evolution 54 (1): 59-75.
Credits
Page created by Axel L. Schönhofer. Dave Carlson helped improve the English.
